Представь детей средь пустоты, где зыбки все координаты.
Н.Коржавин.
Закончен синтез полиизопрена.
Мы близко подошли к разгадке гена.
Но может кость создать из коллагена одна
Природа мудрая пока…Дж.Апдайк. Танец твердых тел.
Роль метафор в научном знании. Особенности метафор генетики
Какие общенаучные и житейские ассоциации, мысли и чувства, возникавшие у различных генетиков при исследованиях новых загадочных объектов и решениях новых необычных проблем, отображались в их профессиональном языке? Имеет ли «коллективное бессознательное» генетиков свои специфические черты и можно ли исследовать пути его развития, изучая словесные образы в публикациях по генетике? Что может дать такой анализ для понимания истории развития самой генетики как системы понятий? И, наконец, что может дать такой анализ генетики для понимания того, что есть знание, наука вообще, и каковы глубинные процессы ее развития?
Оказывается, важнейшие средства научного творчества и вполне рациональной интерпретации новых знаний – это визуальные (изобразительные) и вербальные (словесные) метафоры. Данная работа посвящена лишь вербальным метафорам; анализ визуальных – иная весьма сложная задача.
Что такое научные метафоры вообще и почему они так нужны в различных науках?
Смысл греческого сложного слова «метафора» разъясняют его корни. И в данной работе оно использовано в этом исконном, классическом смысле – в духе Аристотеля. Вербальной научной метафорой будем называть сложное слово или словосочетание, отдельные слова или части слов которого внесены в ту или иную науку из лексиконов других наук, техники и прочих сфер деятельности, где они уже имели иной смысл – специфический, хотя и в чем-то сходный с новым. Это сходство образно и ассоциативно, и поэтому научные метафоры, извлеченные из контекстов, неотличимы от художественных. (Сравним, например, художественные образы «груз воспоминаний» и «бегущее время» с генетическими терминами «генетический груз» и «прыгающие гены»). Среди метафор есть как краткие профессиональные термины – одно сложное слово или сочетание из 2–3 слов, – так и развернутые высказывания – от части фразы до абзаца или совокупности близких по смыслу фраз и их частей в разных частях одного текста.
В англоязычной методологической традиции, к сожалению, почти не известной отечественным читателям, значимость роли метафор в науке обсуждается с начала 1960-х годов [3]. Вот лишь некоторые из ее оценок: «…метафора – это жизненный дух парадигмы (или, точнее, ее основная организующая связь)» [4]; «…любая научная теория – это сеть метафор, и, более того, любое знание, включая научное, неизбежно метафорично» [5]; «метафоры …часто связывают уже сложившиеся концепции с еще только формирующейся новой системой представлений», «метафора выполняет в научном познании организующую функцию, связывая как различные слои языка теории, так и разные по природе и происхождению фрагменты знания» (см. [6], с. 119–133). А сейчас стремительно растет «интерес к метафоре со стороны теории познания, логики, когнитивной психологии, языкознания» (там же, с. 4): начаты исследования метафорических терминов из физики элементарных частиц, астрономии, математики ([6], с. 127) и, наконец, из биологии ХХ века в целом [7]. Однако до сих пор анализ метафор был ограничен рамками гуманитарного дискурса – не было еще ни точных методов анализа, ни структурно-функциональных моделей. Такие новые подходы предложены и обсуждаются ниже.
Профессиональная компетенция автора данной статьи ограничена биологией и, более конкретно, генетикой. Поэтому он не осмеливается анализировать смыслы метафор других наук, по-видимому, тоже представляющие немалый интерес. Метафоры генетики не исследованы вообще. Вероятно, дело в том, что генетикам не до методологической рефлексии – так они поглощены анализом структур и функций самих генов и генных систем, информация о которых лавинообразно растет. А гуманитариев останавливает сложность понятийного аппарата современной генетики.
Более двадцати лет сопоставляя наиболее значительные публикации о структурах и функциях различных генетических систем, автор обнаружил, что именно метафоры – это сама суть формулировок новых фундаментальных подходов, выводов и гипотез. Именно посредством неожиданных и точных образов-словосочетаний наиболее выдающиеся генетики «конструировали» новые, непривычные образы и понятия из уже доступных и употребляемых в других сферах. Ниже мы покажем, что созданные ими тексты могут стать интереснейшим полем исследований для лингвистов, филологов, когнитологов, психологов, методологов и философов. Обычно генетики лишены профессионального снобизма и готовы разъяснять концепции своей науки во благо совместных творческих междисциплинарных исследований.
Как «работают» метафоры в научных концепциях? Вот ряд наших собственных соображений.
Обычно одна и та же сложная природная система – структура или процесс, не целиком изобретенная и созданная человеком, а лишь частично исследованная и понятая им, может быть представленной в умах и в трудах разных исследователей в виде разных систем, состоящих из вербальных и визуальных образов, временных и логических связей, так называемых «когнитивных» моделей. Их изучает когнитивная психология, а в будущем, возможно, они станут объектами нейробиологии и computer science. Разные модели одного явления или же их фрагменты могут быть взаимоисключающими или же частично совместимыми, содержать консенсусы (кстати, в генетике консенсусами называют совпадающие и обобщенные части генетических текстов). Например, в разных моделях одного и того же явления одни и те же факторы могут считаться его причинами или же его следствиями. Понимать и сравнивать такие «разночтения и состыковки» фрагментов моделей необходимо при анализе большинства конкретных научных проблем. Узловые роли в когнитивных моделях играют именно метафоры, неся огромные смысловые нагрузки [8, 9], потому что инновации в лексике и в стилистике совершенно необходимы для выражения новых понятий и формулировок новых концепций. В когнитивной модели того или иного сложного явления метафора играет такую же роль, какую в компьютерной программе играет команда вызова подпрограммы-модуля: она вызывает когнитивный блокассоциацию извне, из другой области знаний, в которой образы, аналогичные исследуемым, уже есть (возможно, такой области соответствует другая зона или нейронная «подсеть» мозга). Поэтому метафору можно рассматривать как своеобразный «концентрат» информации, как новый информационный вход в данную когнитивную модель из других. Конечно, такой прием оказывается очень нужным для описаний реального, воображаемого и предполагаемого не только в живописи, кино и художественной литературе, но и в научных теориях и гипотезах.
Что такое метафоры с позиций математической теории информации? Доказано, что если редкий, неожиданный элемент может быть встроен в систему, не нарушая ее законов, то он может резко повысить ее информационную емкость. Новая метафора – это редкое, неожиданное сочетание слов и смыслов. Поэтому, если она удачна, то в описании увеличивается количество информации (для сравнительно простых систем с известными вероятностями событий вычисляемое по К.Шеннону [10]). Увеличивается и ее ценность, понимаемая как мера «не избыточности, незаменимости информации», для таких простых систем тоже вычисляемая и тем большая, чем выше «уровни рецепции информации» [11]. Потому она максимальна именно на «верхних» уровнях мышления, в описаниях когнитивных моделей сложных процессов. По Г.Кастлеру «создание новой информации состоит в запоминании случайного выбора» [12]. Значит, она создается и при создании метафоры: в процессе авторского творчества происходит перебор и выбор сочетаний морфем и слов, а запоминание удачных вариантов возможно на двух стадиях – автором в его собственных высказываниях и во впервые предлагаемых им терминах и его коллегами в цитируемых высказываниях и в терминах, вошедших в употребление.
Подобную ассоциативную роль играют в живых организмах их важнейшие информационные структуры. Так, гены ключевых этапов развития организмов – это те из «слов» в ДНК (если нуклеотиды сравнивать с буквами), каждое из которых имеет по несколько смыслов: считывание, т.е. работа такого гена на ранних стадиях развития, как бы «связывает в единый букет» несколько весьма различных «сюжетов» – процессов формообразования организма, размножения и дифференцировки его клеток. Таковы многие гомеозисные гены, онкогены и гены мембранных рецепторов. Другой пример на другом структурном уровне – так называемые «гностические» нейроны мозга: каждый из них сопрягает несколько сложных нейронных сетей, формируя ассоциативные структуры – поля и «суперсети» – и этим интегрируя элементарные акты восприятия в образы, а образы – в воспоминания и когнитивные модели. Если сравнить тексты с организмами, то такие гены и такие клетки – это как бы «метафоры во плоти», появившиеся задолго до человеческих языков, речи и письменности.
Биологические и, в частности, генетические системы особенно сложны многоаспектностью, многомасштабностью и иерархичностью структур, функций и отношений сходства и родства. Часто для того или иного конкретного жизненного процесса структурные, функциональные и классификационные схемы не совпадают и представлены различными моделями. А в разных когнитивных моделях того или иного процесса, нередко определяющих его или же определяемых им, считаются разные структурные уровни и даже разные их сочетания. Вот лишь два важных конкретных примера – теории старения и теории рака.
В геронтологии существует несколько сотен (!) не вытекающих друг из друга теорий старения организмов. Самый нижний уровень, предположительно «ведущий» старение, – это нуклеотиды в генах: их мутации портят ферменты, а в итоге – узловые звенья обмена веществ (это «теория катастрофы ошибок»). Следующий уровень – отдельные специальные гены. Например, сейчас усиленно изучают системы, связанные с геном p53: его продукт специфически убивает определенные группы клеток, начиная это делать с раннего зародышевого развития, но особенно активизируясь при старении. В других теориях старение определяется более высокими уровнями: нарушениями общей устойчивости обмена веществ, иммунной и/или эндокринной систем, транспортных систем организма, структур и функций коры головного мозга [13, 14].
Подобно этому, многоуровневыми оказались концепции онкологии. Ведь первичные причины рака – это события, происходящие на разных структурных уровнях. Уровень замен нуклеотидов в генетических текстах – это та же «катастрофа ошибок». Ею может быть мутация в том или ином онкогене (их в геноме человека известно уже более сотни), вызывающая его запоздалую и неумеренную работу – появление неправильного вредоносного белка. Рак могут вызывать мутации, повреждающие системы, каждая из которых состоит из многих белков (и потому кодируется многими генами) и отвечает за тот или иной важнейший клеточный процесс: репарацию ДНК, различные стадии деления клеток, внутриклеточные процессы (их регулируют разнообразные ферменты – протеинкиназы и фосфорилазы), межклеточные взаимодействия (их осуществляют многообразные сигнальные пептиды и мембранные белки-рецепторы), программируемую смерть клеток (это системы, связанные с упомянутым выше геном р53). Онкогенные факторы на уровне систем целого организма, связь которых с перечисленными генетическими системами пока не выяснена, – это нарушения обмена, питания, иммунной системы, эндокринной системы, образа жизни, целостной гармонии организма и даже личностных ценностей [15].
Не менее многообразны и противоречивы различные модели эволюции организмов [16], их эмбрионального развития [17, 18], формирования и существования экологических систем [19] и других важнейших биологических систем и процессов. Каждое из этих явлений описывается несколькими моделями, «опирающимися» на разные структурные уровни и даже на разные их сочетания. Для таких ситуаций философские понятия «холизм» и «редукционизм» необходимы, но далеко не достаточны: чтобы разобраться в современном понимании того или иного сложного и не совсем изученного явления, надо сопоставить различные многоуровневые модели разных авторов. Как будет показано ниже, анализ метафорической лексики – терминов и высказываний – позволяет исследовать важнейшие звенья таких моделей, даже не очень углубляясь в частные эмпирические факты генетики [21, 22].
Важно и то, что внутренний мир генетиков следует понять не только им самим. Ведь их роль в современном миропонимании и деятельности трудно переоценить: они изучают важнейшие принципы размножения и развития разнообразных организмов (включая человека), многие стороны их биохимии и физиологии, процессы старения, множество тяжелых недугов (включая диабет и рак), влияние загрязнений окружающей среды на наследственность, генный контроль биохимических процессов, используемых в пищевой, фармакологической и многих других отраслях промышленности, не говоря уже о таких «бесполезных» (с меркантильных позиций) сюжетах, как принципы и пути эволюции организмов и история жизни на Земле.
Генетика – одна из немногих фундаментальных биологических наук, которая с самого своего зарождения была точной. И вся история ее развития – это история все более и более точных методов и результатов; по способности управлять своими объектами и конструировать их генетика становится все более похожей на физику, математику и инженерно-технические дисциплины. Многочисленные хромосомные и генные карты, записи генетических текстов, схемы строения, работы и эволюции генов и управляемых ими систем и процессов – все эти данные получены точными методами и сами не менее точны, чем алгоритмы и технические чертежи. И потому на первый взгляд ее профессиональный язык – лексика, стилистика и визуальные формы представления данных так далеки от красоты живой природы, открывающейся натуралистам, художникам и поэтам! «Творческий разум осилил – убил…». А если для генетиков понимание красоты живого сводится к анализу разъятых конструкций, к логике и практике решения рациональных задач, то не способны ли они, «как бы резвяся и играя», вообще уничтожить жизнь на Земле, как физик в фантастической притче-романе К.Воннегута «Колыбель для кошки»? И вот, к сожалению, в массовом сознании генетики-экспериментаторы нередко ассоциируются с адептами-жрецами, готовыми из любопытства или корысти прокладывать «пути в незнаемое», не менее чудовищные, чем то, что делал в своих лабораториях Иосиф Менгеле с сотрудниками. Чего стоит новый ажиотаж публики, вызванный клонированием животных и трансплантациями их органов!
А ведь обычно одни и те же генетические знания и методы, а часто и объекты, могут служить для создания как биологического оружия, так и продуктов питания и лекарств. Да, когда генная инженерия еще только зарождалась, именно ее создатели поставили вопросы о возможной генетической опасности, мерах техники безопасности и моральной ответственности генетиков; это стало темой знаменитой конференции в Асиломаре в 1975 году и многократно обсуждалось впоследствии. Но всегда ли исследователи столь нравственны и ответственны? Можно ли утверждать, что исследовательские задачи лежат вне морали и этики, или надеяться, что с развитием точных знаний растет и нравственность? Ведь технократическая цивилизация породила глобальный экологический кризис, к теории естественного отбора апеллировала практика геноцида, расщепленный атом сгубил Хиросиму и Нагасаки, его мирное использование обернулось трагедией Чернобыля… До сих пор биотехнологии приносили гораздо больше благ, чем бед. Но будет ли так и впредь? Что же такое «игры с ДНК» – путь к панацее и «эликсиру жизни» или все же «подарок дьявола» и «мина замедленного действия»? Что сулят обывателю «белые одежды»? Поэтому очень важно было бы знать, что же происходит в умах и чувствах генетиков, когда они вникают в тайны жизни, «алгеброй гармонию поверив».
Такие тревоги могут возникать у школьников, преподавателей биологии и студентов, изучающих генетику лишь как сухую «историю идей» без «истории людей», черпающих дополнительную информацию лишь из прессы, фантастических фильмов и проповедей и впервые листающих специальные статьи. И при этом «за кадром» для них останутся многие сюжеты из прошлого генетики, в которых романтические поиски истины приводили к настоящим пророчествам.
Именно сочетание точности и логической строгости анализа с творческим предвидением структур и функций незримых объектов задолго до их визуализации характерно для работ всех великих генетиков: Г.Менделя, А.Вейсмана, Т.Моргана, Ф.Лежена, Дж.Уотсона и Ф.Крика, Ф.Жакоба и Ж.Моно и многих других. Вся история генетики – это своеобразный «путь вглубь генетических систем», причем «путь впотьмах», на котором нередко исследователи работали «умственными взорами»: в скудном свете новых, порою весьма фрагментарных, фактов они создавали гипотетические схемы и описания тех структур и процессов, которые удавалось опровергать или доказывать экспериментальными данными лишь многие годы спустя. Меры и числа для генетического мышления необходимы, но не достаточны: «провидческие» построения сложных пространственных и временных картин требуют от исследователей не только логики, но и творческого воображения, качества, присущего художникам, писателям и поэтам.
Рассмотрим, как именно это качество запечатлено в лексике и стиле наиболее новаторских трудов генетиков. Нами было замечено, что в них именно неожиданность словосочетаний – основа четкости понятий. На этом наблюдении базируется описанный ниже анализ метафор, характеризующих разные структурные уровни генетических систем. Его логические принципы близки к лингвистическим и наукометрическим методам (в частности, к методам контент-анализа, к тезаурусному и сленговому методам) [1], а также к аналитическим методам молекулярной генетики (двух- и трехмерного электрофореза, дотблот-анализа и т.п.) [2]. Излагаемые ниже результаты показывают, как в профессиональных текстах генетиков, словно в своеобразном магическом кристалле, непроизвольно отображались, фокусировались и сменялись не только рациональные пути развития самой генетики, но и «моды» на различные области точных, естественных и гуманитарных наук, а также житейской практики, в разные периоды нашего века порождавшие разные осознанные и подсознательные мотивы в научном мышлении. Надеемся, что предлагаемые принципы анализа и результаты этой работы будут интересны не только логикам, методологам и философам различных областей науки, а также лингвистам, филологам и психологам, но и самим генетикам-аналитикам, теоретикам и экспериментаторам. Ведь этот подход к истории языка генетики позволяет по-новому сопоставлять не только особенности восприятия и мышления, характерные для разных генетиков и разных периодов генетики, но и сами фундаментальные идеи генетики – разные концепции организации многоуровневых генетических систем. Возможно, этот подход будет полезен и для других эмпирических наук.
Анализ совокупности метафорических терминов
Из глоссария [20] были выписаны все метафорические термины. Их оказалось 859, что составляет около 17% всех терминов глоссария. Следует подчеркнуть, что подавляющее большинство их активно используются в современной генетике. Авторы многих таких метафор – крупнейшие биологи (А.Вейсман, Н.Вавилов, К.Уоддингтон и многие другие); нередко это нобелевские лауреаты (Ф.Крик, Ф.Жакоб, Ж.Моно и др.), и зачастую эти метафоры впервые появлялись именно в работах, удостоенных Нобелевских премий.
Объем данной статьи не позволяет привести полностью список этих терминов и его разбиений на подгруппы. Ниже приведены лишь некоторые из них. Поэтому приведенные ниже количественные результаты и выводы, полученные для всего списка метафорических терминов, а иллюстрируемые лишь некоторыми из них, могут показаться голословными. Однако обсуждаемые здесь результаты проверить легче, чем любой генетический эксперимент – следует лишь повторить работу самим по предлагаемой здесь методике. Для этого достаточно, даже не вникая глубоко в смысл терминов, лишь самим выявить метафоры по «черным словам» глоссария [20] и вновь рассортировать их по предлагаемым ниже критериям. Уверяем, что, несмотря на возможные отдельные расхождения между автором и читателями в классификации некоторых терминов, в целом описанные ниже количественные соотношения воспроизводимы.
Каждый из метафорических терминов был классифицирован по 2 или по 3 критериям:
- 1) из лексикона какой области науки и практики слово (реже – часть слова), входящее в термин, было заимствовано генетиками;
- 2) какой структурный уровень генетических систем им охарактеризован;
- 3) когда он впервые появился (это сделано лишь для тех 677 из 859 метафорических терминов, для которых в глоссарии упомянуты их авторы и даты их создания).
По первым 2 критериям для этой совокупности терминов выявлено:
- – 10 областей заимствования лексики (перечисленных примерно в порядке возрастания сложности описываемых ими систем): физика (с 2 подобластями – «классической» и «субатомной»), химия, география, кибернетика, лингвистика (включая понятия из книгопечатания, относящиеся к текстам), быт, биология, антропология + психология, социология + экономика, мистика;
- – 9 структурных уровней генетических систем (в порядке от больших объектов к малым, т.е. от «верхних» структурных уровней к «нижним»): популяции, семьи + генетические линии, организмы (фенотипы), геномы, хромосомы, плазмиды (включая векторы) и другие столь же малые геномы, системы неаллельных генов, гены как целостные объекты, генетические тексты (логически или явно соответствующие нуклеотидным последовательностям).
В таблице 1 показано распределение метафорических терминов по обоим этим критериям (в ячейках таблицы приведены количества соответствующих терминов). Столбцам соответствуют области заимствования лексики, строкам – генетические структурные уровни, выявленные a posteriori из анализа всего глоссария. Термины, которые можно отнести сразу к нескольким ячейкам, учитывались для каждой из них.
Возможно возражение, что физико-химические характеристики для «нижних» структурных уровней (растворение ДНК, гидролиз полинуклеотидов и т.п.) и анимизирующие для «верхних» (рост популяций, выживание семей и т.п.) – это не метафоры и не термины, а очевидные понятия, и поэтому приведенные соотношения метафор не отражают лексики в целом. Однако анализ количеств таких словосочетаний, проведенный автором по предметным указателям монографий и биологических реферативных журналов, показал, что это не так: если бы учитывались и эти понятия, то в численном отношении преобладали бы те же ячейки, что и в таблице 1, так как в «очевидные» ячейки добавилось бы не более чем по 20–30 выражений.
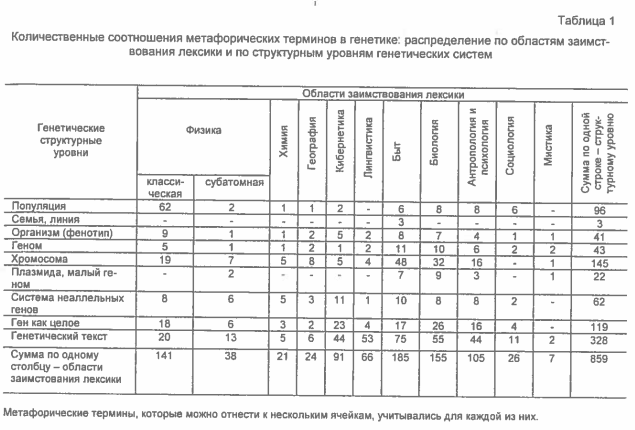
Таблица 1 позволяет сделать несколько выводов.
- Известно, что генетика развивалась, «погружаясь» в исследования все более «нижних» структурных уровней. Соотношения метафорических терминов разных уровней показывают, что этот «путь» противоречил известному методологическому принципу «бритвы Оккама» – «не умножать сущностей». Метафорических терминов, для понимания которых нужны разъяснения, подобные статьям глоссария [20], становилось все больше, а язык генетики – все «эзотеричнее».
- Среди метафорических терминов преобладают:
- – для популяций – физикалистские («вес признака», «генетический груз», «генетическое равновесие», «генный поток», «давление отбора», «квантовое видообразование», «мутационный спектр», «популяционная интерференция», «популяционные волны», «центробежный и центростремительный отбор» и т.п.);
- – для хромосом – бытовые («барабанные палочки», «митотическое веретено», «метафазная пластинка», «полярная шапочка», «репликационная вилка», «стадия букета», «хромосомный мост», «хромосомы типа ламповых щеток» и т.п.) и «анимизирующие» – био и антропоморфные («инициация хромосом», «мобилизация хромосом», «неоцентрическая активность», «сестринские хроматиды», «спаривание хромосом», «хромосомный химеризм», «центросомы» и т.п.);
- – для целых генов и генетических текстов – «анимизирующие» («ассимиляция ДНК», «ген-хозяин», «ген-раб» «гибридизация ДНК», «гнездящиеся (в интронах) гены», «гомеостаз по сплайсингу», «инвазия нитью (ДНК)», «квазивиды», «концертная эволюция», «кочующие (номадические) гены», «миграция ветвей (ДНК)», «молчащая ДНК», «мультигенное семейство», «незаконная рекомбинация», «неразборчивая ДНК», «оператор», «орфон», «отвечающие элементы», «ошибка копирования», «паразитическая ДНК», «популяция последовательностей (ДНК, РНК)», «промотор», «прыгающие гены», «распознавание (кодонов, сайта)», «редактирование (ДНК, РНК)», «сенсорный ген», «созревание РНК», «транспозиционный иммунитет», «употребляемость кодонов», «химерный ген», «чувствительность сайта», «эгоистичная ДНК», «хвост/голова ДНК» и т.п.).
Таким образом, в целом система МТ генетики парадоксальна: совокупности организмов, реально обладающих свободой воли, в ней наиболее уподоблены объектам физики, в то время как гены и ДНК-тексты, изучаемые физико-химическими методами, характеризуются как «живые сущности».
Каковы причины этого?
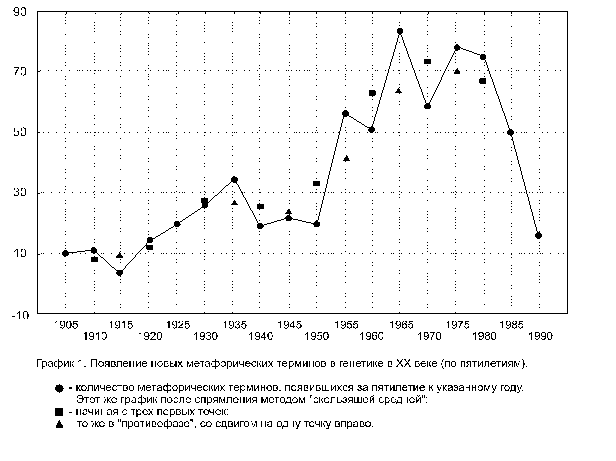
Ситуацию проясняет анализ дат появления различных метафорических терминов.
Для тех 677 из них, которые датированы, была построена кривая их появления – график 1 (ось абсцисс – хронологическая ось, по оси ординат отложено количество терминов, созданных за данный интервал времени).
Перед обеими мировыми войнами, во время и после них на кривой видны прогибы (первый почти нивелируется при спрямлении). Вероятно, генетикам тогда было не до метафор.
Кстати, следует особо отметить, что подобные прогибы, охватывающие периоды этих войн, видны и на хронологических наукометрических графиках других дисциплин, построенных ранее другими авторами на основании анализа количеств докторов наук США, публикаций в физическом реферативном журнале, астрофизических, математических и радиационно-химических исследований, а также научных публикаций в целом (см. [1], с. 224–245), но ими не обсуждавшихся. Возможно, эти прогибы свидетельствуют, что спад различных количественных показателей творчества ученых может служить индикатором и предвестником социальных бедствий и, в частности, больших войн.
Выход на «плато» и снижение кривой в конце графика 1 может отражать либо запаздывание авторов глоссария по отношению к реальному росту количества метафорических терминов, либо предельное «насыщение» ими генетической лексики.
Для сравнительного анализа истории появления различных метафорических терминов были совмещены приемы, использованные при заполнении таблицы 1 и при построении графика 1. Из терминов, относящихся лишь к наиболее полным ячейкам, те из них, которые датированы, были распределены по датам их появления. Таблица 2 является своеобразной комбинацией таблицы 1 и графика 1: ее строкам и подстрокам соответствуют строки и столбцы наиболее полных ячеек таблицы 1, а столбцам – хронологические интервалы.
Видно, что «всплеск физикализма» в создании метафор, терминов генетики популяций, происходил в 1940-е и особенно в 1950–1960-е гг., в период расцвета теоретической и прикладной физики. Метафоры, созданные позже, в основном были характеристиками структурно-функциональных участков геномов: в 1960–1970 гг. – в лингвистико-кибернетическом «типе» тех лет, а после 1970 г. – в «анимизирующем».
Таблица 2
Динамика формирования наиболее многочисленных групп датированных метафорических терминов генетики
|
Генетические структурные уровни |
Области заимствования лексики |
Хронологические интервалы (десятилетия) |
|||||||||
|
|
|
ХIХ в. |
1901 1910 |
1911 1920 |
1921 1930 |
1931 1940 |
1941 1950 |
1951 1960 |
1961 1970 |
1971 1980 |
1981 1990 |
|
Популяция |
биол.+антр.+социол. |
1 |
— |
— |
1 |
1 |
1 |
6* |
1 |
4 |
1 |
|
физика классич. |
— |
2 |
— |
4 |
1 |
7* |
11* |
4 |
2 |
2 |
|
|
Хромосома |
антропология |
— |
1 |
1 |
1 |
— |
1 |
1 |
1 |
2 |
2 |
|
биология |
2 |
1 |
1 |
5 |
4 |
3 |
2 |
3 |
4 |
— |
|
|
быт |
5 |
3 |
— |
1 |
6* |
3 |
2 |
2 |
4 |
4 |
|
|
физика классич. |
1 |
— |
1 |
— |
2 |
1 |
3 |
2 |
— |
— |
|
|
Ген как целое |
антропология |
— |
1 |
1 |
1 |
4 |
— |
1 |
6* |
4 |
6 |
|
биология |
— |
2 |
— |
2 |
2 |
1 |
2 |
3 |
10* |
4 |
|
|
быт |
— |
— |
— |
1 |
1 |
— |
— |
2 |
6 |
2 |
|
|
кибернетика |
— |
— |
— |
— |
— |
— |
3 |
7* |
2 |
2 |
|
|
физика классич. |
— |
— |
1 |
— |
3 |
— |
2 |
3 |
— |
1 |
|
|
антропология |
— |
— |
— |
— |
1 |
— |
3 |
8* |
16* |
7 |
|
|
Генетический текст |
биология |
— |
— |
— |
— |
— |
— |
2 |
7* |
22* |
3 |
|
быт |
— |
— |
— |
1 |
— |
— |
2 |
7* |
15* |
10* |
|
|
лингвистика |
— |
— |
— |
1 |
1 |
— |
2 |
14* |
3 |
1 |
|
|
кибернетика |
— |
— |
— |
— |
— |
— |
4 |
14* |
8* |
5 |
|
|
физика классич. |
— |
— |
— |
— |
— |
— |
1 |
3 |
3 |
— |
|
Cравнительно большие количества помечены (*).
Таким образом, выявлен парадокс: именно тогда, когда генетики стали читать генетические тексты, когда точные методы анализа физической и логической организации генетических систем достигли расцвета и охватили все структурные уровни живого, а количество получаемых ими эмпирических данных об элементарных генных системах экспоненциально росло, в своих образных понятиях генетики стали усиленно интерпретировать эти системы как живые сущности, самостоятельные и даже обладающие свободой воли. Остается неясным, чем это обусловлено: тем ли, что стили восприятия самих генетиков изменялись в соответствии с общими сменами парадигм и эстетических ориентаций в научном сообществе и в обществе в целом, или же тем, что физикалистские, а затем и логико-кибернетические концепции оказывались необходимыми, но не достаточными для понимания объективных особенностей генетических систем.
В обоих случаях, судя по этим особенностям мышления генетиков (по крайней мере авторов терминов), генетике отнюдь не грозит «мертвящий физикализм» восприятия объектов и подходов к ним, чреватый нарушениями биологической и гуманистической этики.
Если каждая «анимизация» нужна лишь затем, чтобы завуалировано охарактеризовать спонтанность, неуправляемость, неконтролируемость и непредсказуемость объекта в условиях исследований, то следует признать, что по мере развития генетики ее терминология все более и более свидетельствовала о непознаваемости генов. Но возможно и другое объяснение: даже генетические тексты реально живут и целесообразно действуют, и потому для их описаний требуется не только технико-физикалистская, но и многообразная биологическая и даже гуманитарная лексика.
В развитии генетики основной вектор исследований был аналитико-редукционистским, направленным «вглубь геномов». Самый «нижний» структурный уровень – конкретные генетические тексты стали доступными анализу двадцать один год назад, с появлением методик их чтения (секвенирования). С тех пор исследованиями охвачены все структурные уровни, и особенно много информации поступило и продолжает поступать о «нижнем» уровне. В генетических текстах прочитаны различные «важнейшие слова», те, от которых зависят ключевые этапы репликации ДНК, размножения клеток, их дифференцировки, формообразования в организмах, злокачественного перерождения, старения… Однако становится все более ясным, что их смыслы проявляются в специфических сложных «контекстах». Один из ярких примеров этого – зависимость проявления гомеозисных генов сегментации тела, HOX II, от стадии и места их проявления и морфологического «контекста» организма.
Стремительное изучение ДНК как «словаря» становится все более ориентированным на изучение тех особенностей генного «синтаксиса», благодаря которым осуществляются целостные «сюжеты». Задачи анализа все более дополняются задачами синтеза, а редукционистское восприятие генных систем – холистическим. По-видимому, усиленная «анимизация» в новых терминах генетики на последнем отрезке ее истории обусловлена именно пониманием сложности и «живости» генетических систем.
Полученные результаты позволяют предположить, что сейчас и в ближайшем будущем развитие генетики будет связано с созданием новых «анимизирующих» метафор: в геномику «нижних», а затем, возможно, и более высоких структурных уровней станут проникать рабочие понятия и модели из экологии, биоценологии, культурологии, психологии, социологии и других биологических и гуманитарных наук, изучающих надорганизменные явления. Насколько мы можем судить по нововведениям 1990–1997 гг. в лексике, синтаксисе и стилистике последних генетических обзоров и устных лекций наших зарубежных коллег, эта тенденция уже весьма выражена.
Будет представлять несомненный интерес поиск в библиографических базах данных новых, еще только формируемых метафор, с помощью запросов типа: ‘(gene OR genes OR genetic) AND » ‘, ‘chromosomе AND » ‘ и т.п.
Подход к анализу метафорических высказываний в проблемных монографиях
Перспективы этого направления работы – моделирование биосистем на базе их словесных описаний, уже созданных биологами. Конкретнее, мы стремимся создать схему эволюции концепций генетики, представив огромный развивающийся массив генетических знаний в виде лексикографических схем, удобных для построения визуальных графических моделей. Такая свертка информации становится все более необходимой теперь, когда ежегодно в мире публикуются буквально сотни тысяч статей с генетической тематикой.
Весьма достойна внимания лингвистов и филологов та литература, которую они до сих пор воспринимают как недоступную их пониманию: базовые проблемные монографии по современной биологии, и в первую очередь по генетике. Кратко покажем, как можно прочитывать эти монографии поновому, даже не обладая фундаментальными знаниями генетики a priori, а приобретая их в минимально необходимых объемах попутно, в таких творческих контактах с генетиками и их литературой, которые как бы нанизывали бы базовые сведения из генетики на стержень лингвистических и филологических исследований.
Острая необходимость в структуралистском диалоге между генетиками и лингвистами назрела вновь, теперь уже в новом контексте, с позиций когнитивной психологии. Для понимания проблем, дискуссий и «белых пятен» самой генетики следует концепции генетических систем, созданные разными авторами, в разных странах и в разные годы, представить в виде когнитивных моделей. Есть надежда, что симбиоз взглядов генетиков, лингвистов, программистов и психологов позволит средствами когнитивной графики строить такие визуальные динамические модели и изучать их поведение, для того чтобы понять, как работают реальные сложные генетические системы. Проверять такие гипотетические модели можно будет в конкретных генетических исследованиях, где сами биосистемы взаимодействуют с логикой и методологией экспериментаторов.
По-видимому, наиболее удобная форма представления таких данных – это блок-схемы, в которых различные генетические структуры и процессы будут представлены в виде узлов и ребер соответствующих графов. Одни из этих объектов обладают «свободой воли» (самопроизвольной динамикой, поведением) и управляют другими, детерминированными извне. Такие свойства биосистем, постулируемые авторами, позволяет выявить их метафорическая лексика и стилистика – анимизация и физикализация объектов в активных и пассивных формах.
Поэтому из всех ячеек, семантических кластеров метафор генетики, описанных выше, теперь мы остановимся лишь на экстремальных, на тех, с помощью которых генетики «анимизировали» или же «физикализировали» свои объекты. С этих позиций рассмотрим метафорические высказывания. Это словосочетания более протяженные, чем термины (от трех слов до абзаца или набора высказываний с общими сюжетами), и, в отличие от терминов, не становящиеся рабочими «инструментами» профессионального сообщества, а сохраняющиеся лишь в авторских текстах. Ограничимся лишь метафорическими высказываниями из нескольких проблемных монографий о структурах и функциях генов и геномов, созданными за последние полвека бурного развития генетики.
Предлагаемая методика вкратце сводится к следующему. Прочитывая монографию, мы извлекаем из нее все «поэтические вольности» – метафорические высказывания автора – и создаем цитатник. Сопоставляя цитаты в рамках такого цитатника, можно строить «метаметафоры» – блок-схемы концептов. Как будет показано ниже, такие модели не только совпадают с резюме (предисловиями, послесловиями и устными оценками) коллеггенетиков, но и выявляют такие важнейшие особенности авторских концепций, которые и сами авторы, и их читатели, и рецензенты-генетики упускают из виду: «за деревьями леса не видно» – обилие эмпирических данных заслоняет логику и «белые пятна» как самих процессов, так и их описаний.
В объеме данной публикации мы сможем лишь остановиться вкратце на нескольких примерах такого анализа, почти не приводя самих метафорических высказываний. Для того чтобы подробно изложить результаты такого анализа, хотя бы только монографий, рассмотренных ниже, необходима целая книга. Поэтому приводимые ниже примеры – лишь «штрихи к портретам» будущих когнитивных моделей. Здесь мы коснемся лишь некоторых выводов, к которым привели сравнения совокупностей метафор в рамках той или иной монографии. Надеемся, что сами эти высказывания – красочные и художественные – станут объектами пристального изучения не только для лингвистов и филологов, но и для специалистов по эстетике. Итак, примеры.
В конце 1960-х гг. М.Ичас в конце своей монографии [23] использовал некоторые метафоры, обсуждая проблемы универсальности и эволюции ДНК-белкового кода, тогда еще только-только расшифрованного: он рассматривал и сам этот код, и его эволюцию как самосовершенствующиеся системы, иногда проявляющие «свободу воли», по-видимому, не сводимые к физико-химическим принципам, однако напоминающие машины.
Событием в изучении молекулярной эволюции (на примере позвоночных животных) стала в 1970–1973 гг. книга С.Оно [24]. В те годы жесткая приверженность дарвинистской парадигме заставила отечественных издателей и переводчиков изменить ее название: оригинальное авторское название "Evolution by Gene Duplication» в советском издании было заменено на «Генетические механизмы прогрессивной эволюции». Предисловие известного генетика Б.Н.Сидорова акцентирует внимание читателей на том, что автор недооценил роль естественного отбора. Однако наш анализ обильных и ярких метафор этой книги показывает: по С.Оно отбор – мощное активное творческое начало, однако в основном судьбу больших групп животных на трудных путях и главных поворотах эволюции, действительно, решали автогенетические процессы – внутренняя логика развития геномов. Построение блок-схемы эволюции генома по С.Оно – задача ближайшего будущего.
Интересный пример бурного развития профессионального дискурса в научной школе – работы В.А.Ратнера и его коллег, более двух десятилетий работающих в Новосибирском Академгородке и развивающих кибернетические методы анализа генетических систем. Сравнивая выводы, сделанные в их монографиях, можно видеть, как мощное развитие кибернетического дискурса, происходившее в 1970-х гг. [25], с середины 1980-х гг. по сей день все активнее дополняется вербальными характеристиками типа «живые свободные гены» [26, 27]. В метафорах, содержащихся в этом цикле работ, гены и генные системы сначала представлялись как механизмы, а затем все более как квазиорганизмы. Эти две парадигмы все более взаимодействуют, причем вербальная анимизация объектов все усиливается.
В начале 1980-х гг., с открытием мобильных генетических элементов («прыгающих генов»), происходила мощная реформация представлений об эволюции геномов. Судя по метафорическим высказываниям в коллективной монографии [28], ее авторов можно разделить на две группы: «редукционистов» (согласно которым пути эволюции геномов определяются самопроизвольными изменениями в отдельных генетических элементах – sensu lato, поведением этих элементов) и «холистов» (согласно которым общая архитектоника геномов – продукт их целостного развития, определяющий эволюционные судьбы отдельных его элементов). «Тот факт, что конференция, по материалам которой была создана эта монография, по сути была диалогом между этими двумя группами генетиков», – констатировал Дж.Мэйнард Смит post factum, закрывая эту конференцию. Это отражено в «Заключении» этой книги.
Р.Б.Хесин в своей монографии, даже название которой имеет несколько анимизирующий метафорический характер [29], сопоставил более 3500 экспериментальных работ, проведенных генетиками мира до 1982 г., и особенно в начале 1980х гг., в последние годы жизни Р.Б.Хесина. Цитатник из этой монографии – свыше 40 развернутых метафорических высказываний – читается как фантастическая сказка. Анимизация в этих высказываниях охватывает лишь один структурный уровень генетической организации – фрагменты ДНК, или генетические элементы. Других метафор нет, так как именно проблеме поведения этих элементов посвящена вся монография. Интересно в нашем контексте следующее.
Возьмем три уровня структурной организации генома: ген (фрагмент ДНК), хромосому и геном. Что чем управляет? Давно известен «холистический» феномен, называемый «эффектом положения гена». Суть этого эффекта в том, что характер работы и проявления гена зависит от того, в какой участок хромосомы этот ген попадает. Эффект положения несколько раз упоминается в [29], однако интересен – и автору, и нам – не этот эффект. В различных местах этой книги метафоры указывают нам на обратные ему, «редукционистские», феномены: спонтанное или индуцированное перемещение некоторых конкретных генетических элементов в другое место генома вызывает его комплексную дестабилизацию – массовые «прыжки» других элементов, множественные хромосомные аберрации и т.п. Этот эффект – когда перемещение элемента внутри системы вызывает ее целесообразную структурную перестройку – интересен и с общефилософских позиций. При этом по метафорической лексике описания таких явлений выявляются в нескольких местах монографии, а подобные феномены описаны и в других ее местах. Полагаем, что, подобно эффекту положения гена, этот генетический метафеномен тоже должен обрести соответствующее названиетермин.
Вступая на путь создания художественных образов (а, как было показано выше, на высоких уровнях научного творчества это неизбежно), ученый, как и поэт, и прозаик, в своих метафорах может и «проговариваться», высказывая отнюдь не то, к чему сам же призывает. Примером этого может служить монография А.Лимаде-Фариа [30]. Стремясь рассматривать биологическую эволюцию, и в первую очередь эволюцию генетических структур, как совокупность чисто физических явлений, для понимания которых достаточно привлекать лишь дисциплины, охватывающие физико-химические феномены, – от квантовой физики до кристаллохимии – автор приводит массу впечатляющих фотографий, иллюстрирующих подобия между живыми и неживыми формами. Однако во многих местах монографии лексика автора выдает прямо противоположное – его скрытый анимизм. Особенно впечатляет глава 17. Не только ее проблематика, но и вся ее лексика и стилистика отражены в ее названии: «Как ген, хромосома и клетка противостоят среде и избегают гибели» (там же, с. 241–258). В частности, по выражению автора, «хромосома… поддерживает постоянство, вводит новшества и производит разведку, пользуясь собственными средствами». Необходимо отметить, что вся эта глава, полная анимизирующих метафор, посвящена именно цитогенетике, полю профессиональных исследований самого автора. Именно в его аналитико-экспериментальных работах ранее были выявлены многие принципы расположения генов в хромосомах и рассмотрена архитектоника хромосомы как целого. Более того, оказывается, что в этой книге, полной метафор, анимизации подвергнуты отнюдь не только биологические объекты. Например, «Элементарные частицы прошли свой эволюционный путь и имеют своих предшественников» (там же, с. 67). И таких примеров в книге масса. Интересно, что эту имплицитную анимизацию живого и неживого на фоне деклараций полного физикализма в биологии не заметил даже глубокий и вдумчивый генетик и философ, редактор русского перевода монографии и автор большого критического послесловия (там же, с. 381–410) Л.И.Корочкин.
Как же понять Лимаде-Фариа? Что это – сознательная подтасовка или скрытая недобросовестность? На наш взгляд, ни то, ни другое. Здесь мы имеем дело со случаем «псевдосубституции» (по терминологии М.Поланьи), ситуацией, когда отрицаемое понятие скрыто вводится в обсуждение как его необходимый элемент. В результате оказывается, что, образно выражаясь, «краеугольные камни здания» теоретических выкладок состоят как бы из концептуального «антивещества», однако «аннигиляции» не происходит: в целом, «здание» концептустойчиво (возможно, лишь до поры до времени).
Следует отметить, что в русскоязычной генетической литературе особой метафоричностью, по-видимому, отражающей свободу творчества, отличаются генетики «новосибирской школы»: Р.Л.Берг, М.Д.Голубовский, В.А.Ратнер и другие. Напротив, в двух обзорных монографиях, изданных в Москве и посвященных тому же кругу проблем, что и в упомянутой монографии С.Оно, – организации и эволюции геномов позвоночных [31, 32] – метафор крайне мало, и все они появляются в конце книг при обсуждении концепций, предлагаемых авторами. Возможно, немногочисленность метафор отражает редакторскую правку или же влияние ее неизбежности на стиль самих этих авторов. Трудно сказать, что это – добросовестность позитивистского описания или же «внутренняя самоцензура». Однако даже основные концепции этих монографий можно выявить, базируясь лишь на этих метафорах. На базе новых молекулярно-генетических фактов оба автора показали, как именно различия в архитектонике хромосом современных позвоночных могут отражать их комплексную реорганизацию, происходившую сотни миллионов лет назад в периоды ароморфозов – выхода на сушу, возникновения зародышевых оболочек, формирования принципов хромосомного определения пола…
Полагаем, что эти несколько примеров уже могут вызвать немалый интерес психологов, лингвистов и филологов, логиков и методологов. Сотрудничая с ними, мы, генетики, сможем поновому понять многочисленные тексты наших коллег, а знатоки, исследователи и ценители языка смогут познать глубины и насущные проблемы генетики и биологии в целом. Надеемся, что предлагаемый нами подход к массиву знаний и проблем, уже сформированному и интенсивно формируемому генетиками, в дальнейшем может позволить увидеть в сжатом и наглядном виде модели развития организма, рака, старения, эволюции разных групп организмов и динамики биологических сообществ. Эти исследования могут быть дополнены и проверены новыми эмпирическими биологическими фактами и экспериментами и осмыслены с помощью невербальных схем и моделей, которые можно будет строить при сравнениях различных биологических публикаций.
Полагаем, что перевод «написанного пером» в мета-вербальные и невербальные формы представления данных, особенно в визуальные динамические модели, станет мощным методологическим инструментом, который обогатит и генетику, и лингвистику, и когнитивную психологию. Возможно, в будущем к этим исследованиям смогут подключиться и специалисты по искусственному интеллекту, моделируя как реальные феномены природы, так и творческие процессы, их интерпретирующие, и нейробиологи, когда они смогут изучать динамику сложных форм знаний в целостном мозге. Таким образом, фактологическая и концептуальная «почва» для совершенно новых междисциплинарных работ сформировалась и ждет своих исследователей.
-
Хайтун С.Д. Наукометрия. Состояние и перспективы. – Москва: Наука, 1983. – С. 122–150.
-
Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование. (Пер. с англ.). – Москва: Мир, 1984.
-
Black M. Models and Metaphors. Ithaca: Cornell University Press, 1962.
-
Haraway D.J. Crystals, Fabrics, and Fields; Metaphors of Organicism in Twentieth-Century Biology. – New Haven: Yale University Press, 1976.
-
Wilden A. System and Structure. Essays in Communication and Exchange. – 2nd ed. – New York: Tavistock Publications, 1980.
-
Метафора в языке и тексте. – Москва: Наука, 1988.
-
Fox Keller E. Refiguring Life: Metaphors of Twentieth Century Biology. – New York: Columbia University Press, 1995.
-
Lakoff G., Johnson M. Metaphors We Live By. – Chicago: The University of Chicago Press, 1980.
-
MacCormac E.R. A Cognitive Theory of Metaphor. – Cambridge (Mass.), MIT Press– London, 1985.
-
Шеннон К. Работы по теории информации и кибернетике. (Пер. с англ.). – Москва: ИЛ, 1963.
-
Волькенштейн М.В. Энтропия и информация. – Москва: Наука, 1986.
-
Кастлер Г. Возникновение биологической организации. (Пер. с англ.). – Москва: Мир, 1967.
-
Finch C.F. Longevity, Senescence and the Genome. – Chicago: The University of Chicago Press, 1991.
-
Rose R. Evolutionary Biology of Aging. – Oxford: Oxford University Press, 1991.
-
Varmus H., Weinberg R.A. Genes and the Biology of Cancer. – New York: Scientific American Library, 1992.
-
Denton M. Evolution: Theory in Crisis. USA, Bethesda, Maryland: Adler & Adler Publishers, Inc., 1986.
-
Гилберт С. Биология развития. (Пер. с англ.). – Москва: Мир, 1994.
-
Goodwin B.C. How the Leopard Changes its Spots. (The Evolution of Complexity). – London: Weidenfeld & Nicolson, 1994.
-
Peters R.H. A Critique for Ecology. Cambridge: Cambridge University Press, 1991.
-
Rieger R., Michaelis A., Green M.M. Glossary of Genetics, Classical and Molecular. – 5th ed. Springer-Verlag, Berlin а.о., 1991.
-
Седов А.Е. История генетики, запечатленная в метафорах ее языка: количественный и структурный анализ. I. Общие принципы анализа. Мета-форические термины // Проблемы социолингвистики и многоязычия. – Москва: Московский Лицей, 1997. – С. 35–47.
-
Седов А.Е. Задачи и принципы анализа метафорических высказываний в проблемных монографиях // Функциональные исследования по лингвистике. – Вып. 6. – Москва: Московский Лицей, 1998. – С. 20–26.
-
Ичас М. Биологический код. – Москва: Мир, 1971. – 351 с.
-
Оно С. Генетические механизмы прогрессивной эволюции. – Москва: Мир, 1973. – 227 с.
-
Ратнер В.А. Молекулярно-генетические системы управления. – Новосибирск: Наука, 1975. – 287 с.
-
Ратнер В.А., Жарких А.А., Колчанов Н.А., Родин С.Н., Соловьев В.В., Шамин В.В. Проблемы теории молекулярной эволюции. – Новосибирск: Наука, 1985. –263 с.
-
Ratner V.A., Zharkikh A.A., Kolchanov N.A., Rodin S.N., Soloviov V.V., Shamin V.V., Antonov A.S. Molecular Evolution. (Biomathematics. Vol. 24). Springer-Verlag: Berlin, Heidelberg, New York, 1996. – 433 p.
-
Эволюция генома / Под ред. Г.Доувера и Р.Флейвелла. – Москва: Мир, 1986. – 368 с.
-
Хесин Р.Б. Непостоянство генома. – Москва: Наука, 1984. – 472 с.
-
Лимаде-Фариа А. Эволюция без отбора. Автоэволюция формы и функции. – Москва: Мир, 1991. – 455 с.
-
Гинатулин А.А. Структура, организация и эволюция генома позвоночных. – Москва: Наука, 1984. – 293 с.
-
Бирштейн В.Я. Цитогенетические и молекулярные аспекты эволюции позвоночных. – Москва: Наука, 1987. – 284 с.
А.Е.Седов, д.б.н., в.н.с.,
Институт истории естествознания
и техники им. С.И.Вавилова РАН
